A. Reproduksi Sel
Pernahkah kalian memikirkan proses tumbuhnya badan bayi hing-ga dewasa?
Dari bayi, kita dapat tumbuh menjadi bentuk sekarang ini disebabkan
sel-sel di dalam tubuh kita terus-menerus memperbanyak diri melalui
pembelahan sel. Oleh karena itu, pembelahan sel meru-pakan faktor
penting dalam hidup kita. Sel merupakan bagian terkecil yang menyusun
tubuh kita. Setiap sel dapat memperbanyak diri dengan membentuk sel-sel
baru melalui proses yang disebut pembelahan sel atau reproduksi sel .
Pada organ-isme bersel satu ( uniseluler ), seperti bakteri dan
protozoa, proses pem-belahan sel merupakan salah satu cara untuk
berkembang biak. Proto-zoa melakukan pembelahan sel dari satu sel
menjadi dua, dari dua sel menjadi empat, dan dari empat sel menjadi
delapan, dan seterusnya.
Pada makhluk hidup bersel banyak (multiseluler), pembelahan sel
mengakibatkan bertambahnya sel-sel tubuh. Oleh karena itu, terjadi-lah
proses pertumbuhan pada makhluk hidup. Pembelahan sel juga berlangsung
pada sel kelamin atau sel gamet yang bertanggung jawab dalam proses
perkawinan antar individu. Setelah dewasa, sel kelenjar kelamin pada
tubuh manusia membelah membentuk sel-sel kelamin.
Seorang laki-laki menghasilkan sperma di dalam testis, sedangkan wanita menghasilkan sel telur atau ovum di dalam ovarium.
Pada dasarnya, pembelahan sel dibedakan menjadi dua, yaitu pembelahan
secara langsung (amitosis) dan pembelahan secara tidak langsung (mitosis
dan meiosis). Apa yang dimaksud dengan pembe-lahan sel secara langsung
maupun tidak langsung tersebut? Kalian akan mengetahuinya dengan
menyimak penjelasan berikut.
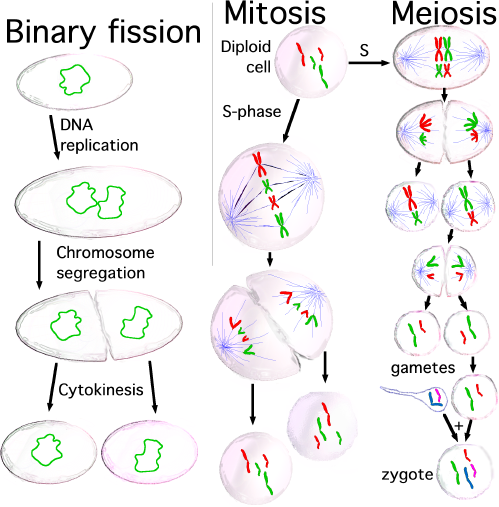
1. Pembelahan Sel secara Langsung
Proses pembelahan secara langsung disebut juga pembelahan ami-tosis atau
pembelahan biner. Pembelahan biner merupakan proses pembelahan dari 1
sel menjadi 2 sel tanpa melalui fase-fase atau tahap-tahap pembelahan
sel. Pembelahan biner banyak dilakukan organisme uniseluler (bersel
satu), seperti bakteri, protozoa, dan mikroalga (alga bersel satu yang
bersifat mikroskopis). Setiap terjadi pembelahan biner, satu sel akan
membelah menjadi dua sel yang identik (sama satu sama lain). Dua sel ini
akan membelah lagi menjadi empat, begitu seterus-nya. Pembelahan biner
dimulai dengan pembelahan inti sel menjadi dua, kemudian diikuti
pembelahan sitoplasma. Akhirnya, sel terbelah menjadi dua sel anakan.
Pembelahan biner dapat terjadi pada organisme prokariotik atau
eukariotik tertentu. Perbedaan antara organisme prokariotik dan
eukariotik, terutama berdasarkan pada ada tidaknya membran inti selnya.
Membran inti sel tersebut membatasi cairan pada inti sel ( nukleoplasma)
dengan cairan di luar inti sel, tempat terdapatnya organel sel (
sitoplasma). Organisme prokariotik tidak mempunyai membran inti sel,
sedangkan organisme eukariotik mempunyai membran inti sel. Oleh karena
itu, eukariotik dikatakan mempunyai inti sel (nukleus) sejati.
Pembelahan biner pada organisme prokariotik terjadi pada bakteri. DNA bakteri terdapat pada daerah yang disebut nukleoid .
DNA pada bakteri relatif lebih kecil dibandingkan dengan DNA pada sel
eukariotik. DNA pada bakteri berbentuk tunggal, panjang dan sirkuler
sehingga tidak perlu dikemas menjadi kromosom sebelum pembelahan.

Contoh organisme eukariotik yang mengalami pembelahan biner adalah
Amoeba. Proses pembelahan sel pada Amoeba dapat kalian pe-lajari pada
Gambar 4.2.
2. Pembelahan Sel secara Tidak Langsung (Mitosis dan Meiosis)
Pembelahan sel secara tidak langsung adalah pembelahan yang melalui
tahapan-tahapan tertentu. Setiap tahapan pembelahan ditandai dengan
penampakan kromosom yang berbeda-beda. Kalian telah mengetahui bahwa di
dalam inti sel terdapat benang-benang kromatin . Ketika sel akan
membelah, benang-benang kromatin ini menebal dan memendek, yang kemudian
disebut kromosom. Kromosom dapat berikatan dengan warna tertentu,
sehingga mudah diamati dengan mikroskop. Hasil penelitian menunjukkan
bahwa kromosom merupakan benang pembawa sifat. Di dalam kromosom
terdapat gen sebagai faktor pembawa sifat keturunan.
Pada waktu sel sedang membelah, terjadi proses pembagian kromosom di
dalamnya. Tingkah laku kromosom selama sel membelah dibedakan menjadi
fase-fase atau tahap-tahap pembelahan sel. Pembelahan sel yang terjadi
melalui fase-fase itulah yang disebut pembelahan secara tidak langsung.
Mengenai fase-fase pembelahan mitosis akan dibahas pada subab
tersendiri.
Pembelahan sel secara tidak langsung dibedakan menjadi dua, yaitu
pembelahan mitosis dan meiosis . Sebelum kalian mempelajari lebih jauh
tentang pembelahan sel secara tidak langsung, ada baiknya kalian lakukan
rubrik Diskusi beri-kut ini.
Proses pertumbuhan dan perkembangan
jaringan atau organ tu-buh organisme terjadi melalui proses pembelahan
sel secara mitosis. Pembelahan mitosis adalah pembelahan sel yang
menghasilkan sel anakan dengan jumlah kromosom sama dengan jumlah
kromosom induknya. Proses pembelahan mitosis terjadi pada semua sel
tubuh makhluk hidup, kecuali pada jaringan yang menghasilkan gamet (sel
kelamin).
Pada pembelahan mitosis, satu sel induk membelah diri menjadi dua sel
anakan. Sel anakan ini mewarisi sifat sel induknya dan memiliki jumlah
kromosom yang sama dengan induknya. Jika sel induk memi-liki 2n
kromosom, maka setiap sel anakan juga emiliki 2n kromo-som. Jumlah 2n
ini disebut juga kromosom diploid .
Pembelahan mitosis terjadi selama pertumbuhan dan reproduksi secara
aseksual. Pada manusia dan hewan, pembelahan mitosis terjadi pada sel
meristem somatik (sel tubuh) muda yang mengalami pertum-buhan dan
perkembangan. Sebagai contoh, sel telur yang telah dibuahi sperma akan
membelah beberapa kali secara mitosis untuk membentuk embrio. Sel-sel
pada embrio ini terus-menerus membelah secara mitosis dan akhirnya
terbentuk bayi. Pertumbuhan manusia dari bayi hingga dewasa juga melalui
mekanisme pembelahan sel secara mitosis. Inilah salah satu bentuk
kekuasaan tuhan yang harus kita syukuri.

Pembelahan meiosis yang disebut juga sebagai pembelahan reduksi
merupakan pembelahan sel induk dengan jumlah kromosom diploid (2n)
menghasilkan empat sel anakan. Setiap sel anakan mengandung separuh
kromosom sel induk atau disebut haploid ( n). Pembelahan meiosis terjadi
pada proses pembentukan sel gamet (sel kelamin) pada organ reproduksi
(testis atau ovarium). Pada manusia atau hewan, sperma yang haploid
dihasilkan di dalam testis dan sel telur yang juga haploid dihasilkan di
dalam ovarium. Pada tumbuhan berbunga, sel gamet dihasilkan di dalam
putik dan benang sari. Pembentukan gamet jantan dan gamet betina terjadi
melalui tahapan gametogenensis (dibahas pada subbab tersendiri).
Penyatuan kedua gamet akan menghasilkan zigot dengan variasi genetik.
Ini disebabkan karena sel anakan merupakan hasil penyatuan dua sel yang
berbeda materi genetiknya. Perpaduan ini menyebabkan adanya variasi
genetik.

B. Tahapan Pembelahan Mitosis
Pembelahan sel secara mitosis meliputi sejumlah tahapan tertentu.
Sebenarnya, pembelahan mitosis hanyalah sebagian kecil dari siklus sel.
Siklus sel terdiri dari fase pembelahan mitosis (M) dan periode
pertumbuhan yang disebut interfase. Interfase merupakan bagian ter-besar
dari siklus sel. Interfase terdiri dari tiga sub fase, yaitu fase G1
(pertumbuhan primer), fase S (sintesis) , dan fase G2 (pertumbuhan
sekunder ).
Pembelahan mitosis merupakan pembelahan yang menghasil-kan sel-sel tubuh (sel somatik). Secara garis besar, pembelahan sel
secara mitosis terdiri dari fase istirahat (interfase ), fase
pembelahaninti sel ( kariokinesis ), dan fase pembelahan sitoplasma
(sitokinesis). Bagaimanakah ciri-ciri setiap fase pembelahan tersebut?
Untuk menge-tahuinya, simaklah penjelasan berikut.
1. Interfase (Fase Istirahat)
Pada tahap ini, sel dianggap sedang istirahat dan tidak melaku-kan
pembelahan. Namun, interfase merupakan tahap yang penting untuk
mempersiapkan pembelahan atau melakukan metabolisme sel. Pada interfase,
tingkah laku kromosom tidak tampak karena berbentuk benang-benang
kromatin yang halus. Walaupun begitu, sel anak yang baru terbentuk sudah
melakukan metabolisme. Sel perlu tumbuh dan melakukan berbagai sintesis
sebelum memasuki proses pembelahan berikutnya.Apa saja kegiatan sel
pada saat interfase? Pada saat interfase, sel mengalami subfase berikut.
a. Fase Pertumbuhan Primer ( Growth 1 disingkat G1 )
Sel yang baru terbentuk mengalami
pertumbuhan tahap pertama. Pada subfase ini, sel-sel belum mengadakan
replikasi DNA yang masih bersifat 2n (diploid). Sementara
organel-organel yang ada di dalam sel, seperti mitokondria, retikulum
endoplasma, kompleks golgi, dan or-ganel lainnya memperbanyak diri guna
menunjang kehidupan sel.
b. Fase Sintesis (S)
Pada subfase ini, sel melakukan sintesis materi genetik. Materi ge-netik
adalah bahan-bahan yang akan diwariskan kepada keturunannya, yaitu DNA.
DNA dalam inti sel mengalami replikasi (penggandaan jumlah salinan).
Jadi, subfase sintesis (penyusunan) menghasilkan 2 salinan DNA.
c. Fase Pertumbuhan Sekunder ( Growth 2 disingkat G2 )
Setelah DNA mengalami replikasi, subfase berikutnya adalah per-tumbuhan
sekunder (G2). Pada subfase ini, sel memperbanyak organel-organel yang
dimilikinya. Ini bertujuan agar organel-organel tersebut dapat
diwariskan kepada setiap sel turunannya. Pada subfase ini, rep-likasi
DNA telah selesai dan sel bersiap-siap mengadakan pembelahan secara
mitosis. Selain itu, inti sel (nukleus) telah terbentuk dengan jelas dan
terbungkus membran inti.
Pada subfase ini, inti sel mempunyai
satu atau lebih nukleolus (membran inti sel). Di luar inti terdapat dua
sentrosom yang terbentuk oleh replikasi sentrosom pada tahap sebelumnya.
Sentrosom mengala-mi perpanjangan menyebar secara radial yang isebut
aster (bintang). Pada sentrosom terdapat sepasang sentriol yang
berfungsi menentukan orientasi pembelahan sel. Walaupun kromosom telah
diduplikasi pada fase S, namun pada fase G2, kromosom belum dapat
dibedakan secara individual karena masih berupa benang-benang kromatin.
Setelah ketiga tahapan interfase
dilalui, sel telah siap menjalani pembelahan secara mitosis. Seperti
fase interfase, pembelahan mitosis juga terdiri dari beberapa fase.
Untuk mengetahui lebih jauh tentang fase-fase pada pembelahan mitosis,
simaklah penjelasan berikut.
2. Pembelahan Mitosis
Kalian telah mengetahui bahwa pembelahan mitosis menghasil-kan sel
anakan yang identik dengan induknya. Secara garis besar, fase pembelahan
mitosis terbagi menjadi dua fase, yaitu fase pembelahan inti (
kariokinesis ) dan fase pembelahan sitoplasma (
sitokinesis).Kariokinesis adalah fase pembelahan inti sel. Secara rinci,
fase kariokinesis dibagi menjadi empat subfase, yaitu profase,
metafase, anafase, dan telofase. Sekarang, marilah kita bahas keempat
subfase tersebut.
a Profase
Pada permulaan profase, di dalam nukleus mulai terbentuk kro-mosom ,
yaitu benang-benang rapat dan padat yang terbentuk akibat menggulungnya
kromatin. Pada fase ini, kromosom dapat dilihat menggunakan mikroskop.
Selanjutnya, nukleolus menghilang dan terjadi duplikasi kromosom
(kromosom membelah dan memanjang) menghasilkan 2 kromosom anakan yang
disebut kromatid . Kedua kromatid tersebut bersifat identik sehingga
disebut kromatid kembar (sister chromatid ), yang bersatu atau
dihubungkan oleh sentromer pada lekukan kromosom. Perhatikan Gambar 4.6.
Sentromer merupakan bagian kromosom yang menyempit, tampak lebih terang
dan membagi kromosom menjadi 2 lengan.
Pada akhir profase, di dalam sitoplasma mulai terbentuk gelendong
pembelahan ( spindel ) yang berasal dari mikrotubulus. Mikrotubulus
tersebut memanjang, seolah-olah mendorong dua sentrosom di sepanjang
permukaan inti sel (nukleus). Akibatnya, sentrosom sa ling menjauh.
Proses ini kemudian berlanjut ke fase berikutnya, yaitu metafase.
b Metafase
Tahap awal metafase (prometafase) ditandai dengan semakin memadatnya kromosom (kromosom ini terdiri dari 2 kromatid) dan
terpecahnya membran inti (membran nukleus). Hal ini menyebab-kan
mikrotubulus dapat menembus inti sel dan melekat pada struktur khusus di
daerah sentromer setiap kromatid, disebut kinetokor . Oleh karena itu,
kinetokor ini berfungsi sebagai tempat bergantung bagi kromosom.
Sebagian mikrotubulus yang melekat pada kinetokor disebut mikro-tubulus
kinetokor, sedangkan mikrotubulus yang tidak memperoleh kinetokor
disebut mikrotubulus non kinetokor. Sementara itu, mikrotubulus non
kinetokor berinteraksi dengan mikrotubulus lain dari kutub sel yang
berlawanan. Pada metafase, kromosom tampak
jelas.
Pada tahap metafase sesungguhnya,
sentrosom telah berada pada kutub sel. Dinding inti sel menghilang.
Sementara itu, kromosom me-nempatkan diri pada bidang pembelahan yang
disebut bidang metafase. Bidang ini merupakan bidang khayal yang
terletak tepat di tengah sel, seperti garis katulistiwa bumi sehingga
disebut juga bidang ekuator. Pada bidang ini, sentromer dari seluruh
kromosom terletak pada satu baris yang tegak lurus dengan gelendong
pembelahan. Kinetokor pada setiap kromatid menghadap pada kutub yang
berlainan (perhatikan Gambar 4.7). Dengan letak kromosom berada di
bidang pembelahan, maka pembagian jumlah informasi DNA yang akan
diberikan kepada sel anakan yang baru, benar-benar rata dan sama
jumlahnya. Tahapan ini merupakan akhir dari metafase.
c Anafase
Setelah berakhirnya tahap metafase, pembelahan sel berlanjut pada tahap anafase. Tahap anafase ditandai dengan berpisahnya
kromatid saudara pada bagian sentromer kromosom. Gerak kromatid ini
disebabkan tarikan benang mikrotubulus yang berasal dari sentriol pada
kutub sel. Kalian telah mengetahui bahwa mikrotubulus melekat pada
sentromer. Hal ini menyebabkan sentromer tertarik terlebih dahulu.
Akibatnya, sentromer berada di depan dan bagian lengan kromatid berada
di belakang. Struktur ini seperti huruf V. Gerakan ini menempuh jarak
sekitar 1μm (10-6 meter) tiap menit. Pada saat bersamaan, mikrotubulus
non kinetokor semakin memanjang sehingga jarak kedua kutub sel semakin
jauh. Selanjutnya, masing-masing kromatid bergerak ke arah kutub yang
berlawanan dan berfungsi sebagai kromosom lengkap, dengan sifat
keturunan yang sama (identik). Untuk menjalankan tugasnya ini,
mikrotubulus telah mengalami peruraian pada bagian kinetokornya. Lalu
bagaimanakah bidang pembelahan sel pada hewan dan tumbuhan?
Salah satu perbedaan sel tumbuhan dan sel hewan adalah ada ti-daknya
sentriol. Pada sel tumbuhan, peran sentriol digantikan oleh kromosom
sehingga arah pembelahan tetap menuju ke kutub sel. Pada sel hewan,
sentriol pada kutub sel merupakan arah yang dituju oleh gerakan kromatid
saat pembelahan.
d Telofase
Pada tahap telofase ini, inti sel anakan terbentuk kembali dari fragmen-fragmen nukleus. Bentuk selnya memanjang akibat peran
mikrotubulus non kinetokor. Benang-benang kromatin mulai longgar. Dengan
demikian, fase kariokinesis yang menghasilkan dua inti sel anak yang
identik secara genetik telah berakhir, namun dua inti sel masih berada
dalam satu sel. Perhatikan Gambar 4.9.
Agar kedua inti terpisah menjadi sel baru, perlu adanya pembelahan sitoplasma yang disebut sitokinesis. Sitokinesis terjadi,
segera setelah telofase selesai. Pada fase sitokinesis terjadi
pembelahan sitoplasma diikuti pembentukan sekat sel baru, sehingga
terbentuk dua sel anakan.
Pada sel hewan, sitokinesis ditandai dengan pembentukan alur pembelahan
melalui pelekukan permukaan sel di sekitar bekas bidang ekuator. Di
sepanjang alur melingkar, terdapat mikrofi lamen yang terdiri dari
protein aktin dan miosin. Protein tersebut berperan dalam kontraksi otot
atau pergerakan sel yang lain. Kontraksi ini semakin ke dalam sehingga
menjepit sel dan membagi isi sel menjadi 2 bagian yang sama.
Berbeda dengan sel hewan, sel tumbuhan mempunyai dinding sel yang keras. Oleh karena itu, pada sitokinensis tidak terbentuk
alur pembelahan. Sitokinesis terjadi dengan pembentukan pelat sel (cell
plate ) yang terbentuk oleh vesikula di sekitar bidang ekuator.
Vesikula-vesikula yang dibentuk oleh badan golgi tersebut saling
bergabung. Penggabungan juga terjadi dengan membran plasma diikuti
terbentuknya dinding sel yang baru oleh materi dinding sel yang dibawa
oleh vesikula.
C. Pembelahan Meiosis
Pernahkah kalian berpikir mengapa seekor kambing hanya mela-hirkan
kambing, manusia melahirkan manusia, atau sapi melahirkan sapi? Secara
kodrati, makhluk hidup tertentu hanya melahirkan makh-luk yang sejenis.
Ini dikarenakan adanya ekanisme tertentu pada saat awal
perkembangbiakan. Bahkan, sebelum terbentuk calon anak di dalam rahim,
mekanisme ini sudah dimulai. Mekanisme ini dimulai pada sel-sel kelamin
(sel reproduksi) calon bapak dan calon ibu. Me-kanisme tersebut adalah
pembelahan sel secara meiosis . Makhluk hidup yang sejenis mempunyai
jumlah kromosom yang sama pada setiap sel. Misalnya, manusia mempunyai
46 kromosom, ke-cuali pada sel reproduksi atau sel kelaminnya. Sel
kelamin pada manusia hanya mempunyai setengah jumlah kromosom sel tubuh
lainnya, yaitu 23 kromosom. Jumlah setengah kromosom (haploid) ini
diperlukan untuk menjaga agar jumlah kromosom anak tetap 46. Kalian
telah mengetahui bahwa anak terbentuk dari perpaduan antara sel kelamin
betina (sel telur) dan sel kelamin jantan (sperma). Perpadu an kedua sel
kelamin yang ma-sing-masing memiliki 23 kromosom ini akan menghasilkan
sel anak (calon janin) yang mempunyai 46 kromosom. Oleh sebab itu,
pembelahan meio-sis sangat berpengaruh dalam perkembang an makhluk
hidup.
Pembelahan meiosis disebut juga
pembelahan reduksi , yaitu pe-ngurangan jumlah kromosom pada sel-sel
kelamin (sel gamet jantan dan sel gamet betina). Sel gamet jantan pada
hewan (mamalia) diben-tuk di dalam testis dan gamet betinanya dibentuk
di dalam ovarium. Gamet jantan pada tumbuhan dibentuk di dalam organ
reproduktif berupa benang sari, sedangkan gamet betinanya dibentuk di
dalam pu-tik. Sel kelamin betina pada hewan berupa sel telur, sedangkan
pada tumbuhan berupa putik. Pada dasarnya, tahap pembelahan meiosis
serupa dengan pembelahan mitosis. Hanya saja, pada meiosis terjadi dua
kali pembelahan, yaitu meiosis I dan meiosis II. Masing-masing
pembelahan meiosis terdiri dari tahap-tahap yang sama, yaitu profase,
metafase, anafase, dan telofase. Bagaimanakah ciri-ciri setiap tahap
pembelahan meiosis tersebut? Kalian akan mengetahuinya setelah
mempelajari uraian berikut.
1. Tahap Meiosis I
Seperti halnya pembelahan mitosis, sebelum mengalami pembe-lahan
meiosis, sel kelamin perlu mempersiapkan diri. Fase persiapan ini
disebut tahap interfase . Pada tahap ini, sel melakukan persiapan berupa
penggandaan DNA dari satu salinan menjadi dua salinan (seperti
interfase pada mitosis). Tingkah laku kromosom masih belum jelas
terlihat karena masih berbentuk benang-benang halus (kro-matin)
sebagaimana interfase pada mitosis. Selain itu, sentrosom juga
bereplikasi menjadi dua (masing-masing dengan 2 sentriol), seperti
tampak pada gambar di samping. Sentriol berperan dalam menentu-kan arah
pembelahan sel.
Setelah terbentuk salinan DNA, barulah
sel mengalami tahap pembelahan meiosis I yang diikuti tahap meiosis II.
Tahap meiosis I ter-diri atas profase I, metafase I, anafase I, dan
telofase I, serta sitokinesis I. Bagaimanakah ciri-ciri setiap fase
pembelahan tersebut? Berikut akan dibahas fase-fase meiosis I pada sel
hewan dengan 4 kromosom diploid (2n = 2). Untuk lebih jelasnya, simaklah
penjelasan di bawah ini.
a. Profase I
Pada tahap meiosis I, profase I merupakan fase terpanjang atau terlama
dibandingkan fase lainnya bahkan lebih lama daripada tahap profase pada
pembelahan mitosis. Profase I dapat berlangsung dalam beberapa hari.
Biasanya, profase I membutuhkan waktu sekitar 90% dari keseluruhan waktu
yang dibutuhkan dalam pembelahan meiosis. Tahapan ini terdiri dari lima
subfase, yaitu leptoten, zigoten, pakiten, iploten, dan diakinesis.
1) Leptoten
Subfase leptoten ditandai adanya benang-benang kromatin yang memendek
dan menebal. Pada subfase ini mulai terbentuk sebagai kromosom homolog.
Kalian perlu membedakan kromosom homolog dengan kromatid saudara. Gambar
4.13 memperlihatkan perbedaan pasangan kromosom homolog dengan kromatid
saudara.
2) Zigoten
Kromosom homolog saling berdekatan atau berpasangan menurut panjangnya. Peristiwa ini disebut sinapsis. Kromosom
homolog yang berpasangan ini disebut bivalen (terdiri dari 2 kro-mosom homolog). Amati kembali Gambar 4.13.
3) Pakiten
Kromatid antara kromosom homolog satu dengan kromosom homolog yang lain disebut sebagai kromatid bukan saudara ( non
sister chromatids ). Dengan demikian, pada setiap kelompok sinap-sis
terdapat 4 kromatid (1 pasang kromatid saudara dan 1 pasang kromatid
bukan saudara). Empat kromatid yang membentuk pa-sangan sinapsis ini
disebut tetrad (Gambar 4.14).
4) Diploten
Setiap bivalen me ngandung empat kromatid yang tetap berkaitan atau berpasangan di suatu titik yang disebut kiasma
(tunggal). Apabila titik-titik perlekatan tersebut lebih dari satu
disebut kiasmata. Proses perlekatan atau persilangan kromatid-kromatid
disebut pindah silang ( crossing over ). Pada proses pin-dah silang,
dimungkinkan terjadinya pertukaran materi genetik
(DNA) dari homolog satu ke homolog lainnya. Pindah silang ini-lah yang memengaruhi variasi genetik sel anakan.
5) Diakinesis
Pada subfase ini terbentuk benang-benang spindel pembela-han (gelendong mikrotubulus). Sementara itu, membran inti sel
atau karioteka dan nukleolus mulai lenyap.Profase I diakhiri dengan
terbentuknya tetrad yang mem-bentuk dua pasang kromosom homolog.
Perhatikan lagi Setelah profase I berakhir, kromosom mulai bergerak ke
bi-dang metafase.
b. Metafase I
Pada metafase I, kromatid hasil duplikasi kromosom homolog berjajar
berhadap-hadapan di sepanjang daerah ekuatorial inti (bidang metafase
I). Membran inti mulai menghilang. Mikrotubulus kinetokor dari salah
satu kutub melekat pada satu kromosom di setiap pasangan. Sementara
mikrotubulus dari kutub berlawanan melekat pada pasang-an homolognya.
Dalam hal ini, kromosom masih bersifat diploid.
c. Anafase I
Setelah tahap metafase I selesai, gelendong mikrotubulus mulai menarik
kromosom homolog sehingga pasangan kromosom homolog terpisah dan
masing-masing menuju ke kutub yang berlawanan Gambar 4.16). Peristiwa
ini mengawali tahap anafase I. Namun, kromatid saudara masih terikat
pada sentromernya dan bergerak sebagai satu unit tunggal. Inilah
perbedaan antara anafase pada mitosis dan meiosis. Pada mitosis,
mikrotubulus memisahkan kromatid yang bergerak ke arah berlawanan. Coba
pelajari lagi tahap anafase pada mitosis.
d. Telofase I
Pada telofase, setiap kromosom homolog telah mencapai kutub-kutub yang
berlawanan. Ini berarti setiap kutub mempunyai satu set kromosom
haploid. Akan tetapi, setiap kromosom tetap mempunyai dua kromatid
kembar. Pada fase ini, membran inti muncul kembali. Peristiwa ini
kemudian diikuti tahap selanjutnya, yaitu sitokinesis.
e. Sitokinesis
Kalian masih ingat pengertian sitokinesis pada sel hewan mau-pun
tumbuhan bukan? Ya, sitokinesis merupakan proses pembelahan sitoplasma.
Tahap sitokinesis terjadi secara simultan dengan telofase. Artinya,
terjadi secara bersama-sama. Tahap ini merupakan tahap di antara dua
pembelahan meiosis. Alur pembelahan atau pelat sel mulai terbentuk
(Gambar 4.17). Pada tahap ini tidak terjadi perbanyakan (replikasi) DNA.
Hasil pembelahan meiosis I menghasilkan dua sel haploid yang
mengandung setengah jumlah kromosom homolog. Meskipun demiki-an,
kromosom tersebut masih berupa kromatid saudara (kandungan DNA-nya masih
rangkap). Untuk menghasilkan sel anakan yang mem-punyai kromosom
haploid diperlukan proses pembelahan selanjutnya, yaitu meiosis II.
Jarak waktu antara meiosis I dengan meiosis II disebut
dengan interkinesis .
Jadi, tujuan meiosis II adalah membagi
kedua salinan DNA pada sel anakan yang baru hasil dari meiosis I.
Meiosis II terjadi pada ta-hap-tahap yang serupa seperti meiosis I. Nah,
untuk mengetahui lebih lanjut tentang tahap meiosis II, perhatikan
uraian selanjutnya.
2. Tahap Meiosis II
Tahap meiosis II juga terdiri dari profase, metafase, anafase, dan
telo-fase. Tahap ini merupakan kelanjutan dari tahap meiosis I.
Masing-masing sel anakan hasil pembelahan meiosis I akan membelah lagi
menjadi dua. Sehingga, ketika pembelahan meiosis telah sempurna,
dihasilkan empat sel anakan. Hal yang perlu diingat adalah bahwa jumlah
kromo-som keempat sel anakan ini tidak lagi diploid (2n) tetapi sudah
haploid
(n). Proses pengurangan jumlah kromosom ini terjadi pada tahap meio-sis
II. Bagaimanakah proses pengurangan jumlah kromosom ini terjadi? Kalian
akan mengetahuinya setelah mempelajari uraian di bawah ini.
a. Profase II
Fase pertama pada tahap pembelahan meiosis II adalah profase II (Gambar
4.18a). Pada fase ini, kromatid saudara pada setiap sel anakan masih
melekat pada sentromer kromosom. Sementara itu, benang mi-krotubulus
mulai terbentuk dan kromosom mulai bergerak ke arah bidang metafase.
Tahap ini terjadi dalam waktu yang singkat karena diikuti tahap
berikutnya.
b. Metafase II
Pada metafase II, setiap kromosom yang berisi dua kromatid, me-rentang
atau berjajar pada bidang metafase II (Gambar 4.18b). Pada tahap ini,
benang-benang spindel (benang mikrotubulus) melekat pada kinetokor
masing-masing kromatid.
c. Anafase II
Fase ini mudah dikenali karena benang spindel mulai menarik kromatid
menuju ke kutub pembelahan yang berlawanan. Akibatnya, kromosom
memisahkan kedua kromatidnya untuk bergerak menuju kutub yang berbeda
(Gambar 4.18c). Kromatid yang terpisah ini se-lanjutnya berfungsi
sebagai kromosom individual.
d. Telofase II
Pada telofase II, kromatid yang telah menjadi kromosom menca-pai kutub
pembelahan. Hasil akhir telofase II adalah terbentuknya 4 sel haploid,
lengkap dengan satu salinan DNA pada inti selnya (nuklei).
e. Sitokinesis II
Selama telofase II, terjadi pula sitokinesis II, ditandai adanya sekat
sel yang memisahkan tiap inti sel. Akhirnya terbentuk 4 sel kembar yang
haploid. Berdasarkan uraian di depan, sel-sel anakan sebagai hasil
pembelahan meiosis mempunyai sifat genetis yang bervariasi satu sama
lain. Variasi genetis yang dibawa sel kelamin orang tua menyebabkan
munculnya keturunan yang bervariasi juga.
D. Gametogenesis dan Pewarisan Sifat
Sebelum menjadi individu baru, baik pada tumbuhan maupun hewan, tentunya
diperlukan bahan baku atau cikal bakal pembentuk in-dividu baru
tersebut. Pada proses perkembangbiakan generatif (seksu-al) hewan maupun
tumbuhan, bahan baku tersebut berupa sel kelamin yang disebut gamet.
Gamet jantan dan betina diperlukan untuk mem-bentuk zigot, embrio,
kemudian individu baru. Nah, pada materi beri-kut ini akan dibahas
tentang proses pembentukan gamet, baik jantan maupun betina yang disebut
gametogenesis ( genesis = pembentukan).Gametogenesis melibatkan
pembelahan meiosis dan terjadi pada organ reproduktif. Pada hewan dan
manusia, gametogenesis terjadi pada testis dan ovarium, sedangkan pada
tumbuhan terjadi pada pu-tik dan benang sari. Hasil gametogenesis adalah
sel-sel kelamin, yaitu gamet jantan (sperma) dan gamet betina (ovum
atau sel telur). Seka-rang, marilah kita mempelajari proses terjadinya
gametoge nesis pada hewan dan tumbuhan.
1. Gametogenesis pada Hewan
Gametogenesis memegang peranan yang
sangat penting dalam perkembangbiakan hewan. Gametogenesis pada hewan
yang akan kita pelajari dibagi menjadi dua, yaitu spermatogenesis dan
oogenesis. Spermatogenesis merupakan proses pembentukan gamet jantan
(sperma).Sementara oogenesis adalah proses pembentuk an gamet betina
(ovum
atau sel telur).
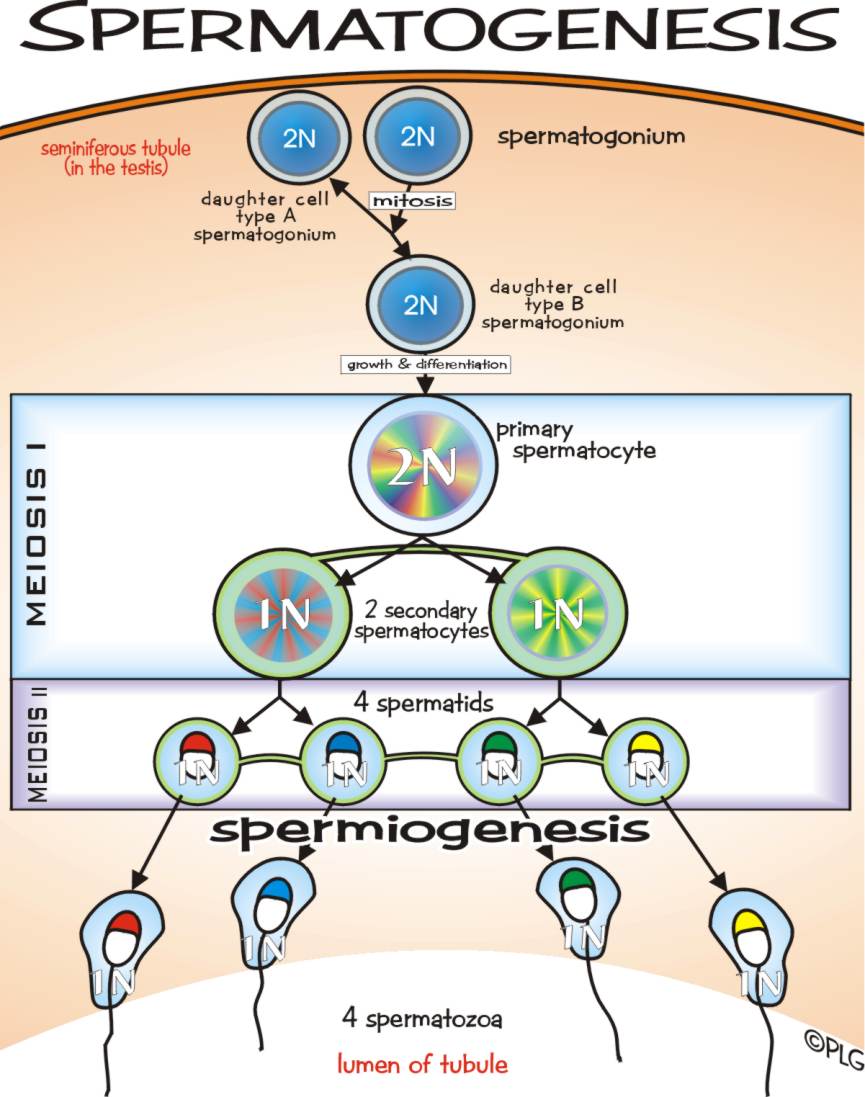
a. Spermatogenesis
Sperma berbentuk kecil, lonjong, berfl agela, dan secara keselu-ruhan
bentuknya menyerupai kecebong (berudu). Flagela pada sperma digunakan
sebagai alat gerak di dalam medium cair. Sperma dihasilkan pada testis.
Pada mamalia, testis terdapat pada hewan jantan sebagai buah pelir atau
buah zakar. Buah pelir pada manusia berjumlah sepasang.
Di dalam testis terdapat saluran-saluran kecil yang disebut tubulus
seminiferus . Pada dinding sebelah dalam saluran inilah, terjadi proses
spermatogenesis. Di bagian tersebut terdapat sel-sel induk sperma yang
bersifat diploid (2n) yang disebut spermatogonium .Pembentukan sperma
terjadi ketika spermatogonium mengalami pembelahan mitosis menjadi
spermatosit primer (sel sperma primer). Selanjutnya, sel spermatosit
primer mengalami meiosis I menjadi dua spermatosit sekunder yang sama
besar dan bersifat haploid. Setiap sel spermatosit sekunder mengalami
meiosis II, sehingga terbentuk 4 sel spermatid yang sama besar dan
bersifat haploid.
Mula-mula, spermatid berbentuk bulat,
lalu sitoplasmanya se-makin banyak berkurang dan tumbuh menjadi sel
spermatozoa yang berfl agela dan dapat bergerak aktif. Berarti, satu
spermatosit primer menghasilkan dua spermatosit sekunder dan akhirnya
terbentuk 4 sel spermatozoa (jamak = spermatozoon ) yang masing-masing
bersifat haploid dan fungsional (dapat hidup).
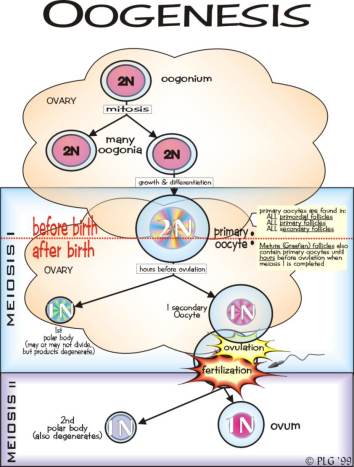
b. Oogenesis
Oogenesis merupakan proses pembentukan sel kelamin betina atau gamet
betina yang disebut sel telur atau ovum. Oogenesis terjadi di dalam
ovarium. Di dalam ovarium, sel induk telur yang disebut oogonium tumbuh
besar sebagai oosit primer sebelum membelah secara meiosis. Berbeda
dengan meiosis I pada spermatogenesis yang menghasilkan 2 spermatosit
sekunder yang sama besar. Meiosis I pada oosit primer menghasilkan 2 sel
dengan komponen sitoplasmik yang berbeda, yaitu 1 sel besar dan 1 sel
kecil. Sel yang besar disebut oosit sekunder , sedangkan sel yang kecil
disebut badan kutub primer ( polar body ).
Oosit sekunder dan badan kutub primer mengalami pembelahan meiosis tahap
II. Oosit sekunder menghasilkan dua sel yang berbeda. Satu sel yang
besar disebut ootid yang akan berkembang menjadi ovum. Sedangkan sel
yang kecil disebut badan kutub.Sementara itu, badan kutub hasil meiosis I
juga membelah menjadi dua badan kutub sekunder. Jadi, hasil akhir
oogenesis adalah satu ovum (sel telur) yang fungsional dan tiga badan
kutub yang me ngalami degenerasi (mati).
Selain pada hewan, gametogenesis juga terjadi pada tumbuhan. Berikut ini akan diuraikan tentang gametogenesis pada tumbuhan
tingkat tinggi.
2. Gametogenesis pada Tumbuhan Tingkat Tinggi
Sebelum menjadi gamet, hasil akhir meiosis pada gametogenesis mengalami
perkembangan terlebih dahulu melalui proses yang dise-but maturasi.
Berikut ini kalian akan membahas proses gametogenesis pada tumbuhan
berbunga (Angiospermae) saja. Pada tumbuhan berbunga, gametogenesis
diperlukan dalam pem-bentukan gamet jantan dan pembentukan gamet betina.
Pembentukan gamet jantan disebut mikrosporogenesis, sedangkan
pembentukan gamet betina disebut megasporogenesis. Mari kita pelajari
pengertian kedua macam gametogenesis tersebut.
a. Mikrosporogenesis
Mikrosporogenesis berlangsung di dalam benang sari, yaitu pada bagian
kepala sari atau anthera . Kepala sari ini meng-hasilkan serbuk sari,
yang mengandung sel sperma. Pembentukan sel sperma dimulai dari sebuah
sel in-duk mikrospora diploid yang disebut mikros porosit di dalam
anthera. Mikrosporosit ini mengalami meiosis I menghasilkan sepasang sel
haploid. Selanjutnya, sel ini mengalami meiosis II dan menghasilkan 4
mikrospora yang haploid. Keempat mikrospora ini berkelompok
menjadi satu sehingga disebut sebagai tetrad .
Setiap mikrospora mengalami pembelahan mi-tosis. Pembelahan ini
menghasilkan dua sel, yaitu sel generatif dan sel vegetatif. Sel vege
tatif ini mempu-nyai ukuran yang lebih besar daripada sel generatif.
Struktur bersel dua ini terbungkus dalam dinding sel yang tebal. Kedua
sel dan dinding sel ini ber-sama-sama membentuk sebuah butiran serbuk
sari yang belum dewasa.
Setelah terbentuk serbuk sari, inti generatif membelah secara mitosis
tanpa disertai sitokinesis, sehingga terbentuklah dua inti sel sperma.
Sementara itu, inti vegetatifnya tidak membelah. Pembentukan sel sperma
ini dapat terjadi sebelum serbuk sari keluardari anthera atau pada saat
serbuk sari sampai di kepala putik (stigma). Pada saat inilah, tangkai
serbuk sari mulai tumbuh. Pada umumnya, pembe-lahan mitosis sel
generatif terjadi setelah buluh serbuk sari menembus stigma atau
mencapai kantung embrio di dalam bakal biji (ovulum).
b. Megasporogenesis
Megasporogenesis merupakan proses pembentukan gamet betina (Gambar 4.24). Proses ini terjadi di dalam bagian betina bunga,
yaitu bakal biji (ovulum) yang dibungkus oleh bakal buah ( ovarium) pada
pangkal putik. Di dalam bakal biji terdapat sporangium yang mengandung
megasporofi t yang bersifat diploid. Selanjutnya, megasporofi t
mengalami meiosis menghasilkan 4 megaspora haploid yang letaknya
berderet. Tiga buah megaspora mengalami degenerasi dan mati, tinggal
sebuah megaspora yang masih hidup.Megaspora yang hidup ini mengalami
pembelahan kromosom secara mitosis 3 kali berturut-turut, tanpa diikuti
pembelahan sitoplasma. Hasilnya berupa sebuah sel besar yang disebut
kandung lembaga muda yang mengan dung delapan inti haploid. Kandung
lembaga ini dikelilingi kulit ( integumen). Di ujungnya terdapat sebuah
lubang (mikropil) sebagai tempat masuknya saluran serbuk sari ke dalam
kandung lembaga.
Selanjutnya, tiga dari delapan inti tadi
menempatkan diri di dekat mikropil. Dua di antara tiga inti yang
merupakan sel sinergid meng-alami degenerasi. Sementara itu, inti yang
ketiga berkembang menjadi sel telur. Tiga buah inti lainnya bergerak ke
arah kutub kalaza , tetapi kemudian mengalami degenerasi pula. Ketiga
inti ini dinamakan inti antipoda . Sisanya, dua inti yang disebut inti
kutub, bersatu di tengah kandung lembaga dan terjadilah sebuah inti
diploid (2n). Inti ini disebut inti kandung lembaga sekunder . Ini
berarti kandung lembaga telah masak, yang disebut megagametofi t dan
siap untuk dibuahi.
3. Pewarisan Sifat dan Variasi Genetis
Secara garis besar, ada tiga mekanisme yang menyebabkan ter-jadinya variasi genetik pada suatu populasi. Ketiga mekanisme ini
dapat dijelaskan sebagai berikut.
a. Pindah silang
Telah dijelaskan di depan bahwa sel kelamin membelah secara meiosis.
Pada profase I, kromosom homolog muncul pertama kali se bagai pasangan.
Kromosom-kromosom homolog ini saling bersilangan pada kiasmata. Pada
kiasmata inilah terjadi pindah silang ( crossing over ) materi genetik
dari kromosom satu ke kromosom lainnya. Pindah silang ini terjadi ketika
dua kromatid dari kromosom yang berbeda bertukar tempat. Kromatid yang
sudah tidak identik lagi dengan kromatid saudaranya karena terjadi
pindah silang disebut dyad . Terjadinya pindah silang ini dapat kalian
lihat pada Gambar 4.24. Pada manusia, dua atau tiga kasus kejadian
pindah silang dapat terjadi untuk setiap pasangan kromosom.
b. Pemilahan kromosom secara bebas
Kalian telah mengetahui bahwa pembelahan sel selalu diikuti pembagian kromosom pada sel anakan yang dihasilkan. Begitu pula
dengan pembelahan meiosis. Pada metafase I, pasangan kromosom homolog
terletak pada bidang metafase. Orientasi pasangan homolog yang menghadap
kutub-kutub sel bersifat acak. Setiap pasangan mempunyai dua
kemungkinan dalam penyusunan ini. Kita ambil contoh organis-me yang
mempunyai empat kromosom diploid (2n = 4). Organisme ini mempunyai 2
kromosom dalam sel gametnya. Dua kromosom ini dapat menghasilkan empat
kemungkinan sel anakan dengan kombinasi kromosom berbeda satu sama lain
Bagaimanakah dengan manusia? Manusia mempunyai 46 kromosom diploid. Ini
berarti pada sperma atau sel telur terdapat 23 kromosom haploid. Dari 23
kromosom ini mempunyai sekitar 8 juta kemungkinan penyusunan homolog
pada metafase. Kandungan kromosom pada sel sperma atau sel telur ini
akan diwariskan pada anak keturunannya. Jadi, setiap manusia sebenarnya
merupakan 1 dari 8 juta kemungkinan pemilahan kromosom yang diwariskan
oleh bapak atau ibu kandungnya.
c. Fertilisasi random
Di dalam sebuah keluarga, seorang anak mempunyai sifat yang berbeda
dengan saudara-saudaranya. Seorang anak tidak ada yang memiliki sifat
yang sama persis dengan ibu atau bapaknya. Akan tetapi, sifatnya
kemungkinan besar merupakan perpaduan sifat kedua orang tuanya. Ini
jelas sangat masuk akal, sebab seorang anak dihasilkan dari pembuahan 1
sel telur ibu oleh 1 sel sperma bapak. Sel telur yang dibuahi sperma
akan menjadi zigot sebagai cikal bakal manusia. Jadi, genetik seorang
anak sangat dipengaruhi kromosom yang terkandung dalam sel telur atau
sperma tersebut. Kalian mengetahui bahwa setiap sel kelamin (sperma dan
sel telur) yang menentukan kromosom anak merupakan 1 dari 8 juta
kemungkinan. Hal ini berarti, seorang manusia merupakan salah satu dari
64 trilyun (8 juta × 8 juta) kombinasi kromosom diploid. Dengan kata
lain, kita telah memenangkan pertandingan melawan 64 trilyun calon anak
yang mungkin dilahirkan. Inilah tanda kebesaran Tuhan Yang Maha Esa.
BAB XV
GENETIKA POPULASI
• Populasi Mendelian
• Frekuensi Genotipe dan Frekuensi Alel
• Polimorfisme Lokus sebagai Indeks Keanekaragaman Genetik
• Hukum Keseimbangan Hardy-Weinberg
• Perubahan Frekuensi Alel
BAB XV. GENETIKA POPULASI
Pola pewarisan suatu sifat tidak selalu dapat dipelajari melalui
percobaan persilangan buatan. Pada tanaman keras atau hewan-hewan dengan
daur hidup panjang seperti gajah, misalnya, suatu persilangan baru akan
memberikan hasil yang dapat dianalisis setelah kurun waktu yang sangat
lama. Demikian pula, untuk mempelajari pola pewarisan sifat tertentu
pada manusia jelas tidak mungkin dilakukan percobaan persilangan. Pola
pewarisan sifat pada organisme-organisme semacam itu harus dianalisis
menggunakan data hasil pengamatan langsung pada populasi yang ada.
Seluk-beluk pewarisan sifat pada tingkat populasi dipelajari pada cabang
genetika yang disebut genetika populasi. Ruang lingkup genetika
populasi secara garis besar oleh beberapa penulis dikatakan terdiri atas
dua bagian, yaitu (1) deduksi prinsip-prinsip Mendel pada tingkat
populasi, dan (2) mekanisme pewarisan sifat kuantitatif. Bagian yang
kedua ini berkaitan dengan penjelasan pada Bab XIV bahwa analisis
genetik sifat-sifat kuantitatif hanya dapat dilakukan pada tingkat
populasi karena individu tidak informatif. Namun, beberapa penulis
lainnya, seperti halnya Bab XV ini, menyebutkan bahwa materi yang
dibahas dalam genetika populasi hanya meliputi deduksi prinsip-prinsip
Mendel pada tingkat populasi.
Populasi dalam arti Genetika
Untuk mempelajari pola pewarisan sifat pada tingkat populasi terlebih
dahulu perlu difahami pengertian populasi dalam arti genetika atau lazim
disebut juga populasi Mendelian. Populasi mendelian ialah sekelompok
individu suatu spesies yang bereproduksi secara seksual, hidup di tempat
tertentu pada saat yang sama, dan di antara mereka terjadi perkawinan
(interbreeding) sehingga masing-masing akan memberikan kontribusi
genetik ke dalam lungkang gen (gene pool), yaitu sekumpulan informasi
genetik yang dibawa oleh semua individu di dalam populasi.
Deskripsi susunan genetik suatu populasi mendelian dapat diperoleh
apabila kita mengetahui macam genotipe yang ada dan juga banyaknya
masing-masing genotipe tersebut. Sebagai contoh, di dalam populasi
tertentu terdapat tiga macam genotipe, yaitu AA, Aa, dan aa. Maka,
proporsi atau persentase genotipe AA, Aa, dan aa akan menggambarkan
susunan genetik populasi tempat mereka berada. Adapun nilai proporsi
atau persentase genotipe tersebut dikenal dengan istilah frekuensi
genotipe. Jadi, frekuensi genotipe dapat dikatakan sebagai proporsi atau
persentase genotipe tertentu di dalam suatu populasi. Dengan perkataan
lain, dapat juga didefinisikan bahwa frekuensi genotipe adalah proporsi
atau persentase individu di dalam suatu populasi yang tergolong ke dalam
genotipe tertentu. Pada contoh di atas jika banyaknya genotipe AA, Aa,
dan aa masing-masing 30, 50, dan 20 individu, maka frekuensi genotipe
AA = 0,30 (30%), Aa = 0,50 (50%), dan aa = 0,20 (20%).
Di samping dengan melihat macam dan jumlah genotipenya, susunan genetik
suatu populasi dapat juga dideskripsi atas dasar keberadaan gennya. Hal
ini karena populasi dalam arti genetika, seperti telah dikatakan di
atas, bukan sekedar kumpulan individu, melainkan kumpulan individu yang
dapat melangsungkan perkawinan sehingga terjadi transmisi gen dari
generasi ke generasi. Dalam proses transmisi ini, genotipe tetua
(parental) akan dibongkar dan dirakit kembali menjadi genotipe
keturunannya melalui segregasi dan rekombinasi gen-gen yang dibawa oleh
tiap gamet yang terbentuk, sementara gen-gen itu sendiri akan mengalami
kesinambungan (kontinyuitas). Dengan demikian, deskripsi susunan genetik
populasi dilihat dari gen-gen yang terdapat di dalamnya sebenarnya
justru lebih bermakna bila dibandingkan dengan tinjauan dari
genotipenya.
Susunan genetik suatu populasi ditinjau dari gen-gen yang ada dinyatakan
sebagai frekuensi gen, atau disebut juga frekuensi alel, yaitu proporsi
atau persentase alel tertentu pada suatu lokus. Jika kita gunakan
contoh perhitungan frekuensi genotipe tersebut di atas, maka frekuensi
alelnya dapat dihitung sebagai berikut.
AA Aa aa Total
Banyaknya individu 30 50 20 100
Banyaknya alel A
60 50 - 110
Banyaknya alel a - 50 40 90
Karena di dalam tiap individu AA terdapat dua buah alel A, maka di dalam
populasi yang mempunyai 30 individu AA terdapat 60 alel A. Demikian
juga, karena tiap individu Aa membawa sebuah alel A, maka populasi yang
mempunyai 50 individu Aa akan membawa 50 alel A. Sementara itu, pada
individu aa dengan sendirinya tidak terdapat alel A, sehingga secara
keseluruhan banyaknya alel A di dalam populasi tersebut adalah 60 + 50 +
0 = 110. Dengan cara yang sama dapat dihitung banyaknya alel a di dalam
populasi, yaitu 0 + 50 + 40 = 90. Oleh karena itu, frekuensi alel A =
110/200 = 0,55 (55%), sedang frekuensi a = 90/200 = 0,45 (45%).
Frekuensi alel berkisar dari 0 hingga 1. Suatu populasi yang mempunyai
alel dengan frekuensi = 1 dikatakan mengalami fiksasi untuk alel
tersebut.
Hubungan matematika antara frekuensi genotipe dan frekuensi alel
Seandainya di dalam suatu populasi terdapat genotipe AA, Aa, dan aa,
masing-masing dengan frekuensi sebesar P, H, dan Q, sementara diketahui
bahwa frekuensi alel A dan a masing-masing adalah p dan q, maka antara
frekuensi genotipe dan frekuensi alel terdapat hubungan matematika
sebagai berikut.
p = P + ½ H dan q = Q + ½ H
Dalam hal ini P + H + Q = 1 dan p + q = 1. Agar diperoleh gambaran yang
lebih jelas mengenai hubungan tersebut, kita perhatikan contoh
perhitungan berikut ini.
Data frekuensi golongan darah sistem MN pada orang Eskimo di Greenland
menurut Mourant (1954) menunjukkan bahwa frekuensi golongan darah M, MN,
dan N masing-masing sebesar 83,5 %, 15,6%, dan 0,9% dari 569 sampel
individu. Kita telah mengetahui pada Bab II bagian alel ganda bahwa
genotipe golongan darah M, MN, dan N masing-masing adalah IMIM, IMIN,
dan ININ. Maka, dari data frekuensi genotipe tersebut dapat dihitung
besarnya frekuensi alel IM dan IN. Frekuensi alel IM = 83,5% + ½ (15,6%)
= 91,3%, sedang frekuensi alel IN = 0,9% + ½ (15,6%) = 8,7%.
Hasil perhitungan frekuensi alel dapat digunakan untuk menentukan sifat
lokus tempat alel tersebut berada. Suatu lokus dikatakan bersifat
polimorfik jika frekuensi alelnya yang terbesar sama atau kurang dari
0,95. Sebaliknya, suatu lokus dikatakan bersifat monomorfik jika
frekuensi alelnya yang terbesar melebihi 0,95. Jadi, pada contoh
golongan darah sistem MN tersebut lokus yang ditempati oleh alel IM dan
IN adalah lokus polimorfik karena frekuensi alel terbesarnya ( IM =
91,3%), masih lebih kecil dari 0,95.
Proporsi lokus polimorfik pada suatu populasi sering kali digunakan
sebagai salah satu indeks keanekaragaman genetik. Nilai lainnya yang
juga sering digunakan sebagai indeks keanekaragaman genetik suatu
populasi adalah heterozigositas rata-rata atau frekuensi heterozigot (H)
rata-rata. Pada contoh di atas besarnya nilai H untuk lokus MN adalah
15,6%. Seandainya dapat diperoleh nilai H untuk lokus-lokus yang lain,
maka dapat dihitung nilai heterozigositas rata-rata pada populasi
tersebut.
Perhitungan frekuensi alel menggunakan data elektroforesis
Frekuensi alel pada suatu populasi spesies organisme dapat dihitung atas
dasar data elektroforesis protein/enzim atau zimogram yang menampilkan
pita-pita sebagai gambaran mobililitas masing-masing polipeptida
penyusun protein (Gambar 15.1). Elektroforesis merupakan teknik
pemisahan molekul yang berbeda-beda ukuran dan muatan listriknya. Oleh
karena itu, molekul-molekul yang akan dipisahkan tersebut harus
bermuatan listrik seperti halnya protein dan DNA.
Jarak
migrasi (cm)
4
3
2
1
Individu 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Genotipe CL LL LL CL CL CL LL CL CL CL LL CL LL LL CL
Gambar 15.1. Zimogram esterase dari ikan sidat (Anguilla sp)
di kawasan Segara Anakan, Cilacap
(Sumber : Susanto, 2003)
Prinsip kerja elektroforesis secara garis besar dapat dijelaskan
sebagai berikut. Sampel ditempatkan pada salah satu ujung media berupa
gel, kemudian kedua ujung gel tersebut diberi aliran listrik selama
beberapa jam sehingga komponen-komponen penyusun sampel akan bergerak
menuju kutub yang muatan listriknya berlawanan dengannya. Kecepatan
gerakan (mobilitas) tiap komponen ini akan berbeda-beda sesuai dengan
ukuran molekulnya. Makin besar ukuran molekul, makin lambat gerakannya.
Akibatnya, dalam satuan waktu yang sama molekul berukuran besar akan
menempuh jarak migrasi yang lebih pendek daripada jarak migrasi molekul
berukuran kecil.
Pola pita seperti pada zimogram esterase di atas sebenarnya merupakan
gambaran fenotipe, bukan genotipe. Namun, analisis variasi fenotipe
terhadap kebanyakan enzim pada berbagai macam organisme sering kali
dapat memberikan dasar genetik secara sederhana. Seperti diketahui, tiap
enzim dapat mengandung sebuah polipeptida atau lebih dengan susunan
asam amino yang berbeda sehingga menghasilkan fenotipe berupa pita-pita
dengan mobilitas yang berbeda. Variasi fenotipe ini disebabkan oleh
perbedaan alel yang menyusun genotipe.
Jika alel-alel yang menyebabkan perbedaan polipeptida pada enzim
tertentu terletak pada suatu lokus, maka bentuk alternatif enzim yang
diekspresikannya dikenal sebagai alozim. Alel yang mengatur alozim
biasanya bersifat kodominan, yang berarti dalam keadaan heterozigot
kedua-duanya akan diekspresikan. Dengan demikian, individu pada Gambar
15.1 yang menampilkan pita lambat dan pita cepat (nomor 1, 4, 5, 6, 8,
9, 10, 12, dan 15) memiliki genotipe heterozigot, yaitu CL (C=cepat;
L=lambat). Sementara itu, individu yang hanya menampilkan pita lambat
(nomor 2, 3, 7, 11, 13, dan 14) adalah homozigot LL. Begitu pula
individu dengan hanya satu pita cepat (kebetulan pada zimogram tersebut
tidak ada) dikatakan mempunyai genotipe homozigot CC.
Dari data genotipe yang diturunkan dari data variasi fenotipe tersebut,
kita dengan mudah dapat menghitung baik frekuensi genotipe maupun
frekuensi alelnya. Frekuensi genotipe CC, CL, dan LL masing-masing
adalah 0, 9/15, dan 6/15. Frekuensi alel C = 0 + ½ (9/15) = 9/30,
sedang frekuensi alel L = 6/15 + ½ (9/15) = 21/30.
Hukum Keseimbangan Hardy-Weinberg
Populasi mendelian yang berukuran besar sangat memungkinkan terjadinya
kawin acak (panmiksia) di antara individu-individu anggotanya. Artinya,
tiap individu memiliki peluang yang sama untuk bertemu dengan individu
lain, baik dengan genotipe yang sama maupun berbeda dengannya. Dengan
adanya sistem kawin acak ini, frekuensi alel akan senantiasa konstan
dari generasi ke generasi. Prinsip ini dirumuskan oleh G.H. Hardy, ahli
matematika dari Inggris, dan W.Weinberg, dokter dari Jerman,. sehingga
selanjutnya dikenal sebagai hukum keseimbangan Hardy-Weinberg.
Di samping kawin acak, ada persyaratan lain yang harus dipenuhi bagi
berlakunya hukum keseimbangan Hardy-Weinberg, yaitu tidak terjadi
migrasi, mutasi, dan seleksi. Dengan perkatan lain, terjadinya
peristiwa-peristiwa ini serta sistem kawin yang tidak acak akan
mengakibatkan perubahan frekuensi alel.
Deduksi terhadap hukum keseimbangan Hardy-Weinberg meliputi tiga
langkah, yaitu (1) dari tetua kepada gamet-gamet yang dihasilkannya, (2)
dari penggabungan gamet-gamet kepada genotipe zigot yang dibentuk, dan
(3) dari genotipe zigot kepada frekuensi alel pada generasi keturunan.
Secara lebih rinci ketiga langkah ini dapat dijelaskan sebagai berikut.
Kembali kita misalkan bahwa pada generasi tetua terdapat genotipe AA,
Aa, dan aa, masing-masing dengan frekuensi P, H, dan Q. Sementara itu,
frekuensi alel A adalah p, sedang frekuensi alel a adalah q. Dari
populasi generasi tetua ini akan dihasilkan dua macam gamet, yaitu A dan
a. Frekuensi gamet A sama dengan frekuensi alel A (p). Begitu juga,
frekuensi gamet a sama dengan frekuensi alel a (q).
Dengan berlangsungnya kawin acak, maka terjadi penggabungan gamet A dan a
secara acak pula. Oleh karena itu, zigot-zigot yang terbentuk akan
memilki frekuensi genotipe sebagai hasil kali frekuensi gamet yang
bergabung. Pada Tabel 15.1 terlihat bahwa tiga macam genotipe zigot akan
terbentuk, yakni AA, Aa, dan aa, masing-masing dengan frekuensi p2,
2pq, dan q2.
Tabel 15.1. Pembentukan zigot pada kawin acak
Gamet-gamet
dan frekuensinya
A
(p) a
(q)
Gamet-gamet
dan frekuensinya A (p)
AA
(p2) Aa
(pq)
a (q) Aa
(pq) aa
(q2)
Oleh karena frekuensi genotipe zigot telah didapatkan, maka frekuensi
alel pada populasi zigot atau populasi generasi keturunan dapat
dihitung. Fekuensi alel A = p2 + ½ (2pq) = p2 + pq = p (p + q) = p.
Frekuensi alel a = q2 + ½ (2pq) = q2 + pq = q (p + q) = q. Dengan
demikian, dapat dilihat bahwa frekuensi alel pada generasi keturunan
sama dengan frekuensi alel pada generasi tetua.
Aplikasi hukum Hardy-Weinberg untuk perhitungan frekuensi alel autosomal
Kemampuan sesesorang untuk merasakan zat kimia feniltiokarbamid (PTC)
disebabkan oleh alel autosomal dominan T. Individu dengan genotipe TT
dan Tt dapat merasakan PTC, sedang individu tt tidak. Pada suatu
pengujian terhadap 228 orang diperoleh bahwa hanya 160 di antaranya yang
dapat merasakan PTC. Dari 160 orang ini dapat dihitung individu yang
bergenotipe TT dan Tt sebagai berikut.
Individu yang tidak dapat merasakan PTC (genotipe tt) jumlahnya 228 –
160 = 68 sehingga frekuensi genotipe tt = 68/228 = 0,30. Dengan mudah
dapat diperoleh frekuensi alel t = √ 0,30 = 0,55 dan frekuensi alel T = 1
– 0,55 = 0,45. Selanjutnya, frekuensi genotipe TT = (0,45)2 = 0,20,
sedang frekuensi genotipe Tt = 2(0,45)(0,55) = 0,50. Banyaknya individu
yang bergenotipe TT = 0,20 x 228 =46, sedang individu yang bergenotipe
Tt = 0,50 x 228 = 114. Jika TT dijumlahkan dengan Tt, maka diperoleh
individu sebanyak 160 orang, yang semuanya dapat merasakan PTC.
Aplikasi hukum Hardy-Weinberg untuk perhitungan frekuensi alel ganda
Salah satu contoh alel ganda yang sering dikemukakan adalah alel
pengatur golongan darah sistem ABO pada manusia. Seperti telah kita
bicarakan pada Bab II, sistem ini diatur oleh tiga buah alel, yaitu IA,
IB, dan I0. Jika frekuensi ketiga alel tersebut masing-masing adalah p,
q, dan r, maka sebaran frekuensi genotipenya = (p + q + r)2 = p2 + 2pq +
2pr + q2 + 2qr + q2. Frekuensi golongan darah A adalah penjumlahan
frekuensi genotipe IA IA dan IA I0 , yakni p2 + 2pr. Demikian pula,
frekuensi golongan darah B, AB, dan O pada suatu populasi dapat dicari
dari sebaran frekuensi tersebut. Sebaliknya, dari data frekuensi
golongan darah (fenotipe) dapat dihitung besarnya frekuensi alel.
Misalnya, dari 500 mahasiswa Fakultas Biologi Unsoed diketahui 196 orang
bergolongan darah A, 73 golongan B, 205 O, dan 26 AB. Alel yang
langsung dapat dihitung frekuensinya adalah I0 , yang merupakan akar
kuadrat frekuensi O. Jadi, frekuensi I0 = √ 205/500 = 0,64. Selanjutnya,
jumlah frekuensi A dan O = p2 + 2pr + r2 = (p + r)2 = (1 – q) 2
sehingga akar kuadrat frekuensi A + O = 1 – q. Dengan demikian,
frekuensi IB (q) = 1 – akar kuadrat frekuensi A + O = 1 –
√(196 + 205)/500 = 0,11. Dengan cara yang sama dapat diperoleh
frekuensi alel IA (p) = 1 – √(73 + 205)/500 = 0,25.
Aplikasi hukum Hardy-Weinberg untuk perhitungan frekuensi alel rangkai X
Telah kita ketahui bahwa pada manusia dan beberapa spesies organisme
lainnya dikenal adanya jenis kelamin homogametik (XX) dan heterogametik
(XY). Pada jenis kelamin homogametik hubungan matematika antara
frekuensi alel yang terdapat pada kromosom X (rangkai X) dan frekuensi
genotipenya mengikuti formula seperti pada autosom. Namun, pada jenis
kelamin heterogametik formula tersebut tidak berlaku karena frekuensi
alel rangkai X benar-benar sama dengan frekuensi genotipe. Pada jenis
kelamin ini tiap individu hanya membawa sebuah alel untuk masing-masing
lokus pada kromosom X-nya. Agar lebih jelas dapat dilihat Tabel 15.2
berikut ini.
Tabel 15.2. Hubungan matematika antara fekuensi alel rangkai X
dan frekuensi genotipe
Homogametik Heterogametik
Genotipe AA Aa aa A a
Frekuensi genotipe P H Q R S
Alel A a A a
Frekuensi alel pm = P + ½H qm = Q + ½H pt = R qt = S
pm = frekuensi alel A pada individu homogametik
qm = frekuensi alel a pada individu homogametik
pt = frekuensi alel A pada individu heterogametik
qt = frekuensi alel a pada individu heterogametik
Untuk seluruh populasi frekuensi alel A dapat dihitung, yaitu p = 2/3 pm
+ 1/3 pt = 1/3 (2 pm + pt) = 1/3 (2P + H + R). Dengan cara yang sama
dapat dihitung pula frekuensi alel a pada seluruh populasi, yaitu q =
2/3 qm + 1/3 qt = 1/3 (2 qm + qt) = 1/3 (2Q + H + S). Kontribusi alel
sebanyak 2/3 bagian oleh individu homogametik disebabkan oleh keberadaan
dua buah kromosom X pada individu tersebut, sementara individu
heterogametik memberikan kontribusi alel 1/3 bagian karena hanya
mempunyai sebuah kromosom X.
Sebagai contoh perhitungan frekuensi alel rangkai X dapat dikemukakan
alel rangkai X yang mengatur warna tortoise shell pada kucing. Misalnya,
dalam suatu populasi terdapat 277 ekor kucing betina berwarna hitam
(BB), 311 kucing jantan hitam (B), 54 betina tortoise shell (Bb), 7
betina kuning (bb), dan 42 jantan kuning (b). Dari data ini dapat
dihitung frekuensi genotipe BB pada populasi kucing betina, yaitu P =
277 / (277+54+7) = 0.82. Sementara itu, frekuensi genotipe Bb (H) = 54 /
(277+54+7) = 0,16 dan frekuensi genotipe bb (Q) = 7 / (277+54+7) =
0,02. Di antara populasi kucing jantan frekuensi genotipe B, yaitu R =
311 / (311+42) = 0,88, sedang frekuensi genotipe b, yaitu S = 42 /
(311+42) = 0,12. Sekarang kita dapat menghitung frekuensi alel B pada
seluruh populasi, yaitu p = 1/3 (2.0,82 + 0,16 + 0,88) = 0,89, dan
frekuensi alel b pada seluruh populasi, yaitu q = 1/3 (2.0,02 + 0,16 +
0,12) = 0,11.
Migrasi
Di atas telah disebutkan bahwa migrasi merupakan salah satu syarat yang
harus dipenuhi bagi berlakunya hukum keseimbangan Hardy-Weinberg. Hal
ini berarti bahwa peristiwa migrasi akan menyebabkan terjadinya
perubahan frekuensi alel. Lebih jauh, kuantifikasi migrasi dalam bentuk
laju migrasi (lazim dilambangkan sebagai m), sering kali digunakan untuk
menjelaskan adanya perbedaan frekuensi alel tertentu di antara berbagai
populasi, misalnya perbedaan frekuensi golongan darah sistem ABO yang
terlihat sangat nyata antara ras yang satu dan lainnya.
Laju migrasi dapat didefinisikan sebagai proporsi atau persentase alel
tertentu di dalam suatu populasi yang digantikan oleh alel migran pada
tiap generasi. Sebagai contoh, jika pada tiap generasi sebanyak 80 dari
1000 ekor ikan normal digantikan oleh ikan albino, maka dikatakan bahwa
laju migrasinya 0,08 atau 8%.
Secara matematika, hubungan antara perubahan frekuensi alel dan laju migrasi dapat dilihat sebagai persamaan berikut ini.
pn – P = (po – P)(1 – m)n
pn = frekuensi alel pada populasi yang diamati setelah n generasi migrasi
P = frekuensi alel pada populasi migran
po = frekuensi alel pada populasi awal (sebelum terjadi migrasi)
m = laju migrasi
n = jumlah generasi
Mutasi
Faktor lain yang dapat menyebabkan terjadinya perubahan frekuensi alel
adalah mutasi. Namun, peristiwa yang sangat mendasari proses evolusi ini
sebenarnya tidak begitu nyata pengaruhnya dalam perubahan frekuensi
alel. Hal ini terutama karena laju mutasi yang umumnya terlalu rendah
untuk dapat menyebabkan terjadinya perubahan frekuensi alel. Selain itu,
individu-individu mutan biasanya mempunyai daya hidup (viabilitas), dan
juga tingkat kesuburan (fertilitas), yang rendah.
Dari kenyataan tersebut di atas dapat dimengerti bahwa mutasi hanya akan
memberikan pengaruh nyata terhadap perubahan frekuensi alel jika mutasi
berlangsung berulang kali (recurrent mutation) dan mutan yang
dihasilkan memiliki kemampuan untuk beradaptasi dengan lingkungan yang
ada.
Hubungan matematika antara laju mutasi dan perubahan frekuensi alel
dapat dirumuskan seperti pada contoh berikut ini. Misalnya, di dalam
suatu populasi terdapat alel A dan a, masing-masing dengan frekuensi
awal po dan qo. Mutasi berlangsung dari A ke a dengan laju mutasi
sebesar u. Sebaliknya, laju mutasi alel a menjadi A adalah v. Dengan
demikian, perubahan frekuensi alel A akibat mutasi adalah ∆p = vqo –
upo, sedang perubahan frekuensi alel a akibat mutasi adalah ∆q = upo –
vqo.
Ketika dicapai keseimbangan di antara kedua arah mutasi tersebut nilai
∆p dan ∆q adalah 0. Oleh karena itu, vqo = upo, atau secara umum vq =
up. Jika persamaan ini dielaborasi, maka akan didapatkan p = v/(u + v)
dan q = u/(u + v).
Seleksi
Sebegitu jauh kita mengasumsikan bahwa semua individu di dalam populasi
akan memberikan kontribusi jumlah keturunan yang sama kepada generasi
berikutnya. Namun, kenyataan yang sebenarnya sering dijumpai tidaklah
demikian. Individu-individu dapat memberikan kontribusi genetik yang
berbeda karena mereka mempunyai daya hidup dan tingkat kesuburan yang
berbeda.
Proporsi atau persentase kontribusi genetik suatu individu kepada
generasi berikutnya dikenal sebagai fitnes relatif atau nilai seleksi
individu tersebut. Nilai fitnes relatif berkisar antara 0 dan 1.
Genotipe superior di dalam suatu populasi, atau disebut juga genotipe
baku, dikatakan memiliki nilai fitnes relatif sama dengan 1, sementara
untuk genotipe-genotipe lainnya nilai fitnes relatif besarnya kurang
dari 1. Proporsi pengurangan kontribusi genetik suatu genotipe bila
dibandingkan dengan kontribusi genetik genotipe baku disebut koefisien
seleksi (s) genotipe tersebut. Dengan perkataan lain, nilai fitnes
relatif genotipe ini adalah 1 – s.
Kembali kita misalkan bahwa di dalam suatu populasi terdapat genotipe
AA, Aa, dan aa. Kondisi dominansi ketiga genotipe ini berdasarkan atas
nilai fitnes relatifnya dapat dilihat pada Gambar 15.2 berikut ini.
aa Aa AA
(1-s) (1-½s) 1
a)
aa Aa AA
(1-s) (1-½s) 1
b)
aa AA/Aa
(1-s) 1
c)
aa AA Aa
(1-s2) (1-s1) 1
d)
Fitnes relatif
Gambar 15.2. Berbagai kondisi dominansi dilihat dari nilai fitnes relatifnya
a) Semi dominansi
b) Dominansi parsial
c) Dominansi penuh
d) Overdominansi
Pada kondisi semi dominansi dan dominansi parsial (Gambar 15.2 a dan b)
genotipe Aa memberikan kontribusi genetik yang lebih kecil bila
dibandingkan dengan kontribusi genotipe baku (AA), sedang pada kondisi
dominansi penuh (Gambar 15.2 c) genotipe ini memberikan kontribusi
genetik sama besar dengan kontribusi genotipe AA. Bahkan pada kondisi
overdominansi, genotipe Aa menjadi genotipe baku dan kontribusi
genetiknya justru lebih besar daripada kontribusi genotipe AA. Dominansi
heterozigot (kondisi overdominansi) ini dapat dijumpai misalnya pada
kasus resistensi individu karier anemia bulan sabit (sickle cell anemia)
terhadap penyakit malaria. Individu dengan genotipe homozigot HbSHbS
akan mengalami pengkristalan molekul hemoglobin, dan eritrositnya
berbentuk seperti bulan sabit, sehingga individu ini akan menderita
anemia berat dan biasanya meninggal pada usia muda. Namun, individu
heterozigot HbSHbA justru memiliki ketahanan yang lebih tinggi terhadap
infeksi parasit penyebab malaria bila dibandingkan dengan individu
normal (HbAHbA). Di tempat-tempat yang menjadi endemi penyakit malaria,
genotipe HbSHbA merupakan genotipe baku (fitnes relatif = 1), sedang
individu normal HbAHbA mempunyai nilai fitnes relatif kurang dari 1.
Perubahan frekuensi alel akibat seleksi berlangsung sesuai dengan
kondisi dominansi yang ada. Pada kondisi dominansi penuh, misalnya,
perubahan frekuensi alel dapat dihitung sebagai berikut.
Genotipe AA Aa aa Total
Frekuensi awal p2 2pq q2 1
Fitnes relatif 1 1 1 – s
Kontribusi genetik p2 2pq q2(1 – s ) 1 – sq2
Terlihat bahwa kontribusi genetik total mejadi lebih kecil dari 1 karena
genotipe aa mempunyai nilai fitnes relatif 1 – s. Dari rumus hubungan
matematika antara frekuensi alel dan frekuensi genotipe dapat dihitung
besarnya frekuensi alel a setelah seleksi, yaitu q1 = q2(1 – s ) + pq
/ 1-sq2. Jika perubahan frekuensi alel a dilambangkan dengan ∆q,
maka ∆q = q1 – q = q2(1 – s ) + pq / 1-sq2 – q. Setelah
persamaan ini kita elaborasi akan didapatkan ∆q = – sq2( 1 – q) / 1 –
sq2. Untuk kondisi dominansi yang lain besarnya perubahan frekuensi alel
akibat seleksi dapat dirumuskan dengan cara seperti di atas.
Sistem Kawin Tidak Acak
Faktor lain yang meyebabkan gangguan keseimbangan Hardy-Weinberg adalah
sistem kawin tidak acak (non random mating). Jika dilihat dari segi
fenotipe, ada sistem kawin tidak acak yang dikenal sebagai perkawinan
asortatif. Dengan perkataan lain, perkawinan asortatif adalah sistem
kawin tidak acak yang didasarkan atas fenotipe.
Perkawinan asortatif dapat berupa perkawinan asortatif positif atau
asortatif negatif (disasortatif). Pada perkawinan asortatif positif
individu-individu yang mempunyai fenotipe sama cenderung untuk lebih
sering bertemu bila dibandingkan dengan individu-individu dengan
fenotipe berbeda. Sebaliknya, pada perkawinan asortatif negatif
individu-individu yang mempunyai fenotipe berbeda cenderung untuk lebih
sering bertemu bila dibandingkan dengan individu-individu dengan
fenotipe yang sama.
Di samping perkawinan asortatif ada pula sistem kawin tidak acak yang
tidak memandang fenotipe individu tetapi dilihat dari hubungan
genetiknya. Sistem kawin semacam ini dapat dibedakan menjadi dua macam,
yaitu silang dalam (inbreeding) dan silang luar (outbreeding). Silang
dalam adalah perkawinan di antara individu-individu yang secara genetik
memiliki hubungan kekerabatan, sedang silang luar adalah perkawinan di
antara individu-individu yang secara genetik tidak memiliki hubungan
kekerabatan. Perkawinan asortatif positif dan silang dalam akan
meningkatkan frekuensi genotipe homozigot. Sebaliknya, perkawinan
asortatif negatif dan silang luar akan meningkatkan frekuensi genotipe
heterozigot.
Silang dalam
Contoh silang dalam yang paling ekstrim dapat dilihat pada tanaman yang
melakukan penyerbukan sendiri. Katakanlah generasi pertama suatu
populasi tanaman menyerbuk sendiri hanya terdiri atas individu-individu
dengan genotipe Aa. Oleh karena terjadi penyerbukan sendiri di antara
genotipe Aa, maka pada generasi kedua dari seluruh populasi akan
terdapat genotipe AA, Aa, dan aa masing-masing sebanyak 1/4, 1/2, dan
1/4 bagian. Pada generasi ketiga genotipe AA dan aa akan bertambah 1/8
bagian yang berasal dari segregasi genotipe Aa pada generasi kedua.
Sebaliknya, genotipe Aa akan berkurang menjadi 1/4 bagian sehingga
populasi generasi ketiga akan terdiri atas (1/4+1/8) AA, 1/4 Aa, dan
(1/4+1/8) aa atau 3/8 AA, 1/4 Aa, 3/8 aa. Dengan demikian, sampai dengan
generasi ketiga saja sudah terlihat bahwa frekuensi genotipe homozigot,
baik AA maupun aa, mengalami peningkatan, sedang frekuensi heterozigot
Aa berkurang.
Genotipe homozigot untuk suatu lokus tertentu – jika kita berbicara
individu normal diploid – mempunyai dua buah alel yang sama pada lokus
tersebut. Persamaan di antara dua alel pada genotipe homozigot dapat
terjadi dengan dua kemungkinan. Pertama, mereka secara fungsional sama
sehingga menghasilkan fenotipe yang sama pula. Dua alel semacam ini
dikatakan sebagai alel serupa (alike in state). Kemungkinan kedua,
mereka berasal dari hasil replikasi sebuah alel pada generasi
sebelumnya. Jika hal ini yang terjadi, maka kedua alel tersebut
dikatakan seasal atau identik (identical by descent).
Untuk menggambarkan besarnya peluang bahwa dua buah alel yang sama pada
individu homozigot merupakan alel identik digunakan suatu nilai yang
disebut sebagai koefisien silang dalam (inbreeding coefficient). Nilai
ini besarnya berkisar dari 0 hingga 1, dan biasanya dilambangkan dengan
F. Nilai F sama dengan 0 apabila kedua alel pada individu homozigot
tidak mempunyai asal- usul yang sama atau merupakan hasil kawin acak.
Sebaliknya, nilai F sama dengan 1 apabila kedua alel sepenuhnya
merupakan alel identik atau berasal dari leluhur bersama (common
ancestor) yang sangat dekat.
Besarnya nilai F dapat dihitung dari diagram silsilah seperti contoh
pada Gambar 15.3. Misalnya, individu A kawin dengan B menghasilkan dua
anak, yaitu C dan D. Selanjutnya, kakak beradik C dan D kawin, mempunyai
anak X. Koefisien silang dalam individu X dapat dihitung sebagai
berikut.
A B * Hitung jumlah
loop. Loop adalah jalan yang menghubungkan kedua orang tua
C D X (C dan D)
melewati leluhur bersama (A dan B). Pada soal ini terdapat dua
X loop, yaitu CAD dan CBD.
Gambar 15.3.Contoh diagram silsilah *Hitung jumlah individu yang terdapat pada tiap loop sebagai nilai n.
* Hitung nilai F dengan rumus :
F = Σ (½)n(1 + FA)
n = jumlah
individu yang terdapat pada tiap loop (pada soal ini terdapat 3
individu, baik pada loop CAD maupun CBD)
FA = koefisien silang dalam leluhur bersama (pada soal ini FA dan FB
masing-masing sama dengan 0 karena dianggap sebagai individu hasil kawin
acak)
Dengan demikian, nilai F individu X (FX) pada contoh soal tersebut di
atas adalah (½)3(1 + 0) + (½)3(1 + 0) = ¼. Hal ini berarti bahwa
peluang bertemunya alel-alel identik yang berasal dari leluhur bersama,
baik A maupun B, pada individu X besarnya ¼.
Makin besar nilai F, makin cepat diperoleh tingkat homozigositas yang
tinggi. Sebagai gambaran, pembuahan sendiri dapat mencapai homozigositas
100% pada generasi keenam, sementara perkawinan antara saudara kandung
baru mencapainya pada generasi keenam belas. Peningkatan homozigositas
akibat silang dalam dapat menimbulkan tekanan silang dalam (inbreeding
depression) apabila di antara alel-alel identik yang bertemu terdapat
sejumlah alel resesif yang kurang menguntungkan.
Perubahan frekuensi alel yang disebabkan oleh terjadinya silang dalam
dapat dihitung dari perubahan frekuensi genotipe seperti pada Tabel
15.3.
Tabel 15.3. Frekuensi genotipe hasil kawin acak
dan silang dalam
Genotipe Frekuensi
Kawin acak Silang dalam
AA p2 p2 (1 – F) + pF
Aa 2 pq 2 pq (1 – F)
aa q2 q2 (1 – F) + qF
Jika nilai F = 0, maka frekuensi genotipe AA, Aa, dan aa masing-masing
adalah p2, 2 pq, dan q2 . Frekuensi tersebut ternyata sama dengan
frekuensi genotipe hasil kawin acak. Jika nilai F = 1, maka frekuensi
genotipe AA, Aa, dan aa masing-masing menjadi p, 0, dan q. Hal ini
berarti di dalam populasi hanya tinggal individu homozigot, sedang
individu heterozigot tidak dijumpai lagi.
Silang luar
Berkebalikan dengan silang dalam, silang luar akan meningkatkan
frekuensi heterozigot. Di samping itu, jika silang dalam dapat
menyebabkan terjadinya tekanan silang dalam yang berpengaruh buruk
terhadap individu yang dihasilkan, silang luar justru dapat memunculkan
individu hibrid dengan sifat-sifat yang lebih baik daripada kedua
tetuanya yang homozigot. Fenomena keunggulan yang diperlihatkan oleh
individu hibrid hasil persilangan dua tetua galur murni (homozigot)
disebut sebagai vigor hibrida atau heterosis.
Ada beberapa teori mengenai mekanisme genetik yang menjelaskan
terjadinya heterosis. Salah satu di antaranya adalah teori dominansi,
yang pada prinsipnya menyebutkan bahwa alel-alel reseif merugikan yang
dibawa oleh masing-masing galur murni akan tertutupi oleh alel dominan
pada individu hibrid yang heterozigot. Misalnya, ada alel A yang
menyebabkan akar tanaman tumbuh kuat sementara alel a menjadikan akar
tanaman lemah. Sementara itu, alel B menyebabkan batang menjadi kokoh,
sedang alel b menyebabkan batang lemah. Persilangan antara galur murni
AAbb (akar kuat, batang lemah) dan aaBB (akar lemah, batang kuat) akan
menghasilkan hibrid AaBb yang mempunyai akar dan batang kuat.
Fenomena heterosis sudah sering sekali dimanfaatkan pada bidang
pemuliaan tanaman, antara lain untuk merakit varietas jagung hibrida.
Galur murni A disilangkan dengan galur murni B, mendapatkan hibrid H.
Namun, karena biji hibrid H ini dibawa oleh tongkol tetuanya (A atau B)
yang kecil, maka jumlah bijinya menjadi sedikit dan tidak cukup untuk
dijual kepada petani. Oleh karena itu, jagung hibrida yang dipasarkan
biasanya bukan hasil silang tunggal (single cross) seperti itu,
melainkan hasil silang tiga arah (three-way cross) atau silang ganda
(double cross). Pada silang tiga arah hibrid H digunakan sebagai tetua
betina untuk disilangkan lagi dengan galur murni lain sehingga biji
hibrid yang dihasilkan akan dibawa oleh tongkol hibrid H yang ukurannya
besar. Agak berbeda dengan silang tiga arah, pada silang ganda hibrid H
disilangkan dengan hibrid I hasil silang tunggal antara galur murni C
dan D. Dalam silang ganda ini, sebagai tetua betina dapat digunakan baik
hibrid H maupun hibrid I karena kedua-duanya mempunyai tongkol yang
besar.







